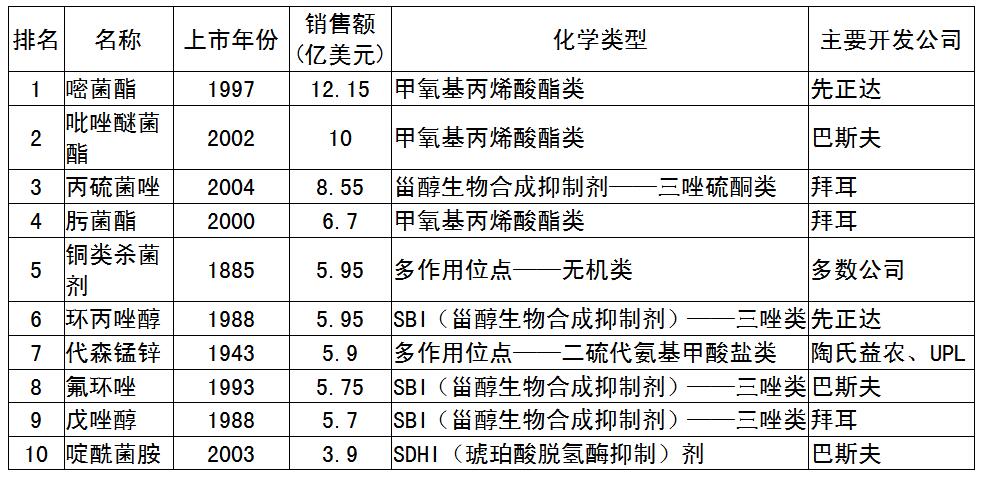
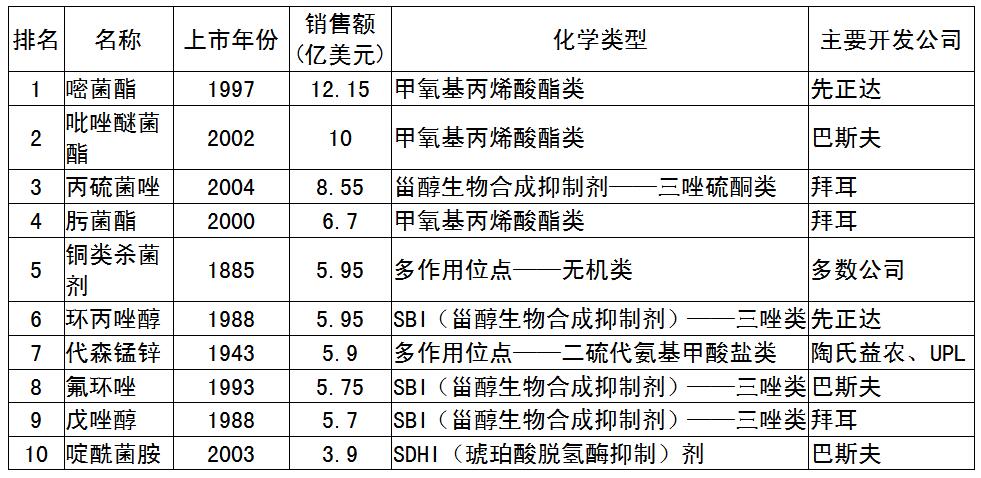
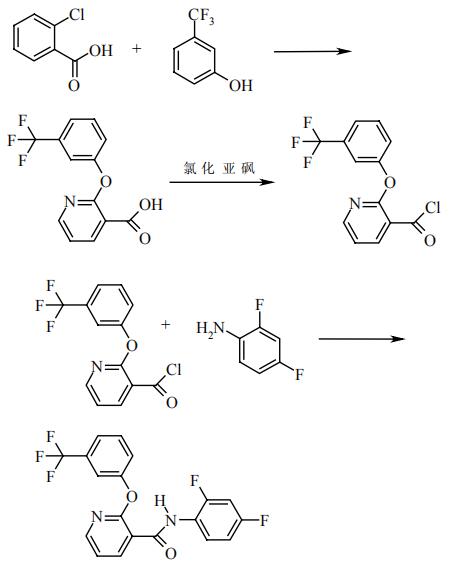
(3)恶苗病的防治
除栽培措施外,水稻种子处理是防治恶苗病最有效的方法。
① 苯并咪唑类杀菌剂。日本在20世纪70年代后用苯菌灵、甲基硫菌灵浸种和拌种防治恶苗病,使用时间长达10年之久。我国在20世纪80年初,应用多菌灵浸种防治恶苗病,取得了较好的防治效果,在东北稻区、江浙沪稻区、河北稻区、长江中游稻区都大面积应用。但在20世纪90年代,由于恶苗病菌对苯并咪唑类杀菌剂产生较高的抗药性而被淘汰。
② DMIs类杀菌剂。在恶苗病菌对多菌灵等苯并咪唑类杀菌剂产生抗药性后,许多研究者筛选其他杀菌剂替代多菌灵,如溴硝醇、乙蒜素等,也在生产上推广应用。DMIs类等三唑类杀菌剂被许多研究者看中,如烯唑醇、三唑醇、种菌唑等,但以咪鲜胺的效果最好,安全性较好,应用面积最大。咪鲜胺在结构上属酰胺类杀菌剂,其作用机理也是抑制甾醇脱氢酶活性。咪鲜胺在20世纪90年代末应用于生产后,在2006年发现了恶苗病菌对咪鲜胺的低敏感菌株,生产上也反映咪鲜胺对恶苗病的防治效果下降。
③ 氰烯菌酯。由江苏省农药研究所创制的氰基丙烯酸酯类杀菌剂,该药对部分镰刀菌具有较高的抑制效果,在小麦赤霉病防治中发挥较大作用。我们的研究表明,其对水稻恶苗病也具有较高的防治效果,可以大面积推广应用。但因为氰基丙烯酸酯类杀菌剂作用位点单一,抗药性风险较高,在应用氰烯菌酯防治水稻恶苗病时,应加强恶苗病菌对其抗药性检测。
④ 其他杀菌剂。二硫氰基甲烷(浸种灵、浸丰)对恶苗病具有较好的防治效果,因为原药生产上的问题而在生产上淘汰。近期的研究表明,氟啶胺、咯菌腈对恶苗病菌具有较高的抑菌效果,用这两种药剂拌种或浸种对恶苗病的防治效果可达90%左右。
(4)恶苗病菌抗药性
水稻恶苗病菌对杀菌剂抗药性研究集中在对多菌灵等苯并咪唑类杀菌剂和对咪鲜胺抗药性研究。
① 对苯并咪唑类杀菌剂抗药性。日本岩手县、滋贺县于20世纪80年代首次报道了对苯菌灵具抗性的恶苗病菌株,高抗菌株(MIC≥100 mg/L)占83.4%。90年代初,我国在连续多年使用多菌灵及复配剂的地区也发现了抗该类药物菌株的存在。我们在1990—1992年分别对从全国4省市共16个县采集的菌株进行测定,结果高抗菌株占65.9%,抗性菌株已成这些地区的优势种群。抗多菌灵菌株与苯菌灵、甲基硫菌灵、噻菌灵等并苯咪唑类杀菌剂均有交互抗药性,且抗性菌株适合度下降不明显。
经过对恶苗病菌对多菌灵具有不同抗性水平菌株的β-微管蛋白基因核苷酸序的比较研究,表明水稻恶苗病菌的β-微管蛋白的165、198、200和257位置氨基酸未发生突变,在克隆的片段内也未发现能引起氨基酸改变的核苷酸突变。说明恶苗病菌对多菌灵产生抗性的分子机理与目前已知抗多菌灵的其他真菌有所不同,有待进一步研究。
② 对咪鲜胺的抗药性。研究表明,水稻恶苗病菌对咪鲜胺的抗药性近年来发展较快,抗性菌株的频率从1.8%上升到67.3%,即在江苏地区咪鲜胺抗性种群已上升为优势种群,而在上海稻区,由于常年使用乙蒜素浸种防治恶苗病,没有发现抗咪鲜胺菌株。
对脱甲基酶CPY51的基因进行PCR扩增和比对,抗性菌株的Cyp51A基因的第13位密码子编码的氨基酸由缬氨酸(Val)变化为甘氨酸(Gly),而这一变化可能是菌株产生抗药性的原因。
水稻恶苗病菌抗咪鲜胺菌株转管培养5~20代后,菌株抗性有所下降,竞争力有所下降。交互抗药性研究结果表明,咪鲜胺与氰烯菌酯、种菌唑、戊唑醇、多菌灵和氟吡菌酰胺之间没有交互抗药性。
③ 对其他杀菌剂的抗性。水稻恶苗病菌对二硫氰基甲烷(浸种灵)的敏感基线为(1.265±0.014) mg/L(EC50),以5 mg/L(MIC)为标准,未检测到抗浸种灵菌株。供试水稻恶苗病菌对氰烯菌酯的EC50分布于0.0317~1.717 mg/L间,平均EC50值为(0.274±0.210) mg/L,未发现敏感性下降的亚群体。
目前恶苗病菌对氰烯菌酯、乙蒜素、二硫氰基甲烷均未发现田间抗性菌株。
2 水稻细菌性病害
我国发生的水稻细菌性病害主要有白叶枯病、细菌性条斑病和细菌性基腐病。
2.1 白叶枯病
水稻白叶枯病是一种世界性的重要细菌病害,而以日本、印度和中国发生比较严重。我国主要发生在华东、华中、华南稻区,在西北、西南、华北和东北部分稻区也有分布。
声明
来源:互联网
本文地址:http://farm.00-net.com/nz/ny/2019/2/20/ny_12603.html